



For optometrists who have been practicing for more than a decade, rods and cones were the only photoreceptors discussed in school. Optometry schools are just now beginning to mention the third photoreceptor in the retina. Similarly, KMK board preparation only mentions the intrinsically photosensitive retinal ganglion cell (ipRGC) one time in a total of 3 books for Part I and Part II.
In the clinical realm, you will be hard-pressed to find optometrists who mention this cell to their patients. After researching this novel photoreceptor for 4 years, I began wondering why the optometry profession as a whole seemingly neglects such a unique and clinically relevant cell. Perhaps it stems from an ignorance of this cell’s significance, from doubts about the ipRGC implications, or even from a lack of education about its existence.
Despite the unpopularity of the ipRGC, this cell assuredly affects both you and your patients throughout the day and even as you sleep. Such knowledge can affect your management of specific patients, contribute to additional glasses revenue, and help you take better care of your patients.
Rods and Cones
Each of you are likely intimately familiar with rods and cones, however, I will provide a brief overview as a reminder as a means to better explain the unique characteristics of ipRGCs.

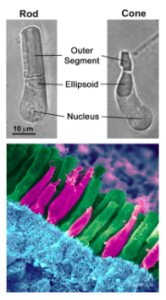
Both rods and cones, now referred to as ‘traditional’ or ‘classical’ photoreceptors, are photoreceptors located in the outer retina, specifically in the photoreceptor layer. These traditional photoreceptors are predominantly involved in image-forming functions and hyperpolarize in response photons.
Rods are primarily found in the peripheral retina and are responsible for scotopic vision. Providing less detailed vision, rods are highly sensitive to dim light, shape, and movement.
Abundant in the central retina are cones, which provide sharp, detailed vision as well as color discrimination.
The Unexplained
For years, rods and cones were believed to be the only photoreceptors in the human retina. However, many visual phenomena were observed which could not be explained through rods and cones alone.
In 1923, Clyde Keeler witnessed strong pupil responses in mice blinded by outer retinal disease. Scientists were also troubled by the fact that some blind humans experienced photophobia while others did not.
Similarly, some blind humans retained regular sleep wake patterns, while others had profound circadian rhythm chronodisruption. More recently, in 1990, Russell Foster genetically engineered mice devoid of rods and cones and was astonished to find normal pupillary light responses and sleep/wake patterns.
Enucleated mice were found to lack normal circadian rhythm, offering further support of a third photoreceptor.
Discovery of the Intrinsically Photosensitive Retinal Ganglion Cell

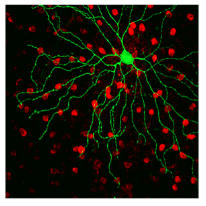
In 1998, Iggy Provencio discovered a novel photopigment never seen before in frog skin. Two years later, this photopigment was found in human retinal ganglion cells and dubbed “melanopsin”.
Soon thereafter, David Berson mechanically isolated these cells containing melanopsin and found them to be capable of detecting light, thus proving that these cells are indeed photoreceptors. These new photoreceptors were given the name, intrinsically photosensitive retinal ganglion cells (ipRGCs).
Anatomy of ipRGCs
ipRGCs have remarkable and unique characteristics, differing them traditional photoreceptors. While traditional photoreceptors are located in the outer retina (photoreceptor layer), ipRGCs are located in the ganglion cell layer of the inner retina.
Obviously, this is significant, as eye care practitioners know some retinal diseases affect the inner retina, while others are known to primarily affect the outer retina. This explains why mice and humans blinded by the outer retinal disease have still demonstrated normal pupil responses.

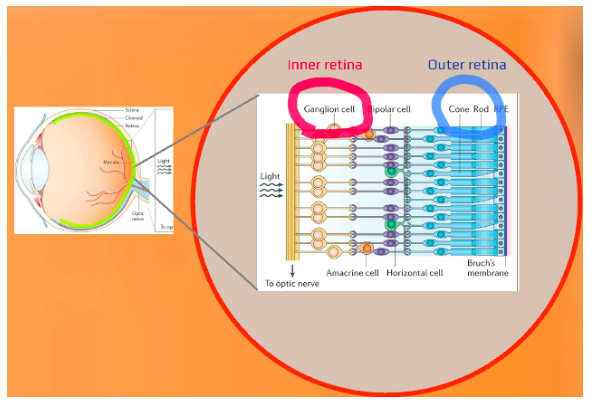
While traditional photoreceptor axons project to the lateral geniculate nucleus (LGN), superior colliculus (SC), and pretectal nucleus (PN), ipRGCs project to not only those three brain structures but dozens more areas as well.
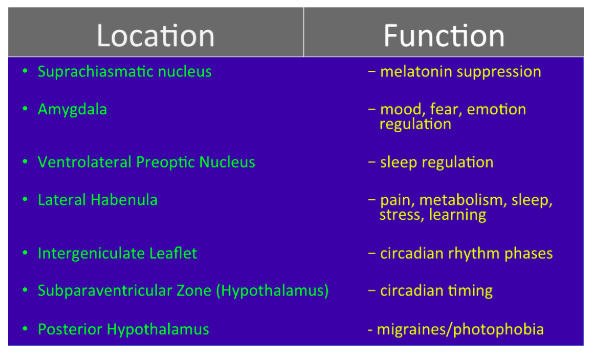
Notable brain areas receiving ipRGC axonal projections include the suprachiasmatic nucleus, amygdala, ventrolateral preoptic nucleus, lateral habenula, intergeniculate leaflet, subparaventricular zone of the hypothalamus, and the posterior hypothalamus.

Physiology of ipRGCs
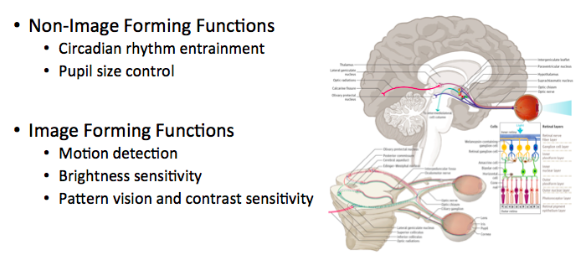
Traditional photoreceptors are almost exclusively involved in image forming roles, the only exception being pupil size control. Conversely, ipRGCs are primarily involved in the non-image forming functions of circadian rhythm photoentrainment and pupil size control.
ipRGC image forming functions include motion detection, brightness sensitivity, and possibly contrast sensitivity and pattern vision.
Research of ipRGCs is still in its infancy, but projections of ipRGCs to dozens of other brain areas give credence to the possibility of ipRGCs playing a role in learning, memory, emotion modulation, fear, mood, pain, and even metabolism.
Early research is beginning to support such implications. What is known for certain is that ipRGCs have undeniable and substantial influences on circadian rhythm timing, sleep/wake regulation, melatonin regulation, photophobia, and migraine exacerbation.

Next Time
In Part II of this series, we will delve into more detail about ipRGC stimulation, ipRGC involvement in photophobia and circadian rhythm, improving sleep quality, blue light effects, and clinical implications of ipRGCs.
